- Leishmania
Taxobox
color = khaki
name = "Leishmania"
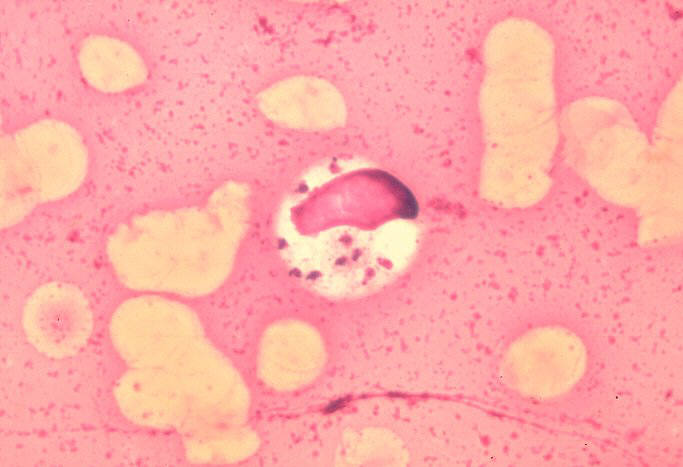
image_width = 240px
image_caption = "Leishmania donovani" in bone marrow cell.
domain = Eukaryota
unranked_phylum =Excavata
phylum =Euglenozoa
classis =Kinetoplastida
ordo =Trypanosomatida
genus = "Leishmania"
subdivision_ranks = Species
subdivision = "L. aethiopica"
"L. amazonensis"
"L. arabica"
"L. archibaldi" (disputed species)
"L. aristedesi"
"L. (Viannia) braziliensis"
"L. chagasi" (syn. "L. infantum")
"L. (Viannia) colombiensis"
"L. deanei"
"L. donovani"
"L. enriettii"
"L. equatorensis"
"L. forattinii"
"L. garnhami"
"L. gerbili"
"L. (Viannia) guyanensis"
"L. herreri"
"L. hertigi"
"L. infantum"
"L. killicki"
"L. (Viannia) lainsoni"
"L. major"
"L. mexicana"
"L. (Viannia) naiffi"
"L. (Viannia) panamensis"
"L. (Viannia) peruviana"
"L. (Viannia) pifanoi"
"L. (Viannia) shawi"
"L. tarentolae"
"L. tropica"
"L. turanica"
"L. venezuelensis""Leishmania" is a
genus oftrypanosome protozoa , and is theparasite responsible for the diseaseleishmaniasis .cite book | author = Ryan KJ; Ray CG (editors) | title = Sherris Medical Microbiology | edition = 4th ed. | pages = 749–54 | publisher = McGraw Hill | year = 2004 | isbn = 0838585299 ] cite book | author = Myler P; Fasel N (editors). | title = Leishmania: After The Genome | publisher = Caister Academic Press | year = 2008 | id = ISBN 978-1-904455-28-8 ] It is spread through sandflies of the genus "Phlebotomus " in theOld World , and of the genus "Lutzomyia " in theNew World . Their primary hosts arevertebrate s; "Leishmania" commonly infectshyrax es,canid s,rodent s, andhuman s. "Leishmania" currently affects 12 million people in 88 countries.Origin
The origins of "Leishmania" are unclear.cite journal | author = Momen H, Cupolillo E | title = Speculations on the origin and evolution of the genus Leishmania | journal = Mem. Inst. Oswaldo Cruz | volume = 95 | issue = 4 | pages = 583–8 | year = 2000 | pmid = 10904419 | doi = 10.1590/S0074-02762000000400023 | accessdate = 2007-05-26] cite journal | author = Noyes HA, Morrison DA, Chance ML, Ellis JT | title = Evidence for a neotropical origin of Leishmania | journal = Mem. Inst. Oswaldo Cruz | volume = 95 | issue = 4 | pages = 575–8 | year = 2000 | pmid = 10904417 | doi = 10.1590/S0074-02762000000400021 | accessdate = 2007-05-26] One possible theory proposes an African origin, with migration to the Americas. Another migration from the Americas to the
Old World about 15 million years ago, across theBering Strait land bridge. Another proposes apalearctic origin.cite journal | author = Kerr SF | title = Palaearctic origin of Leishmania | journal = Mem. Inst. Oswaldo Cruz | volume = 95 | issue = 1 | pages = 75–80 | year = 2000 | pmid = 10656708 | doi = 10.1590/S0074-02762000000100011| accessdate = 2007-05-26] Such migrations would entail migration of vector and reservoir or successive adaptations along the way. A more recent migration is that of "L. infantum" from Mediterranean countries toLatin America (there named "L. chagasi"), since European colonization of theNew World , where the parasites picked up its current New World vectors in their respective ecologies. This is the cause of the epidemics now evident. One recent New World epidemic concerns foxhounds in the USA.Pathophysiology
"Leishmania" cells have two morphological forms: "promastigote" (with an anterior
flagellum ) [http://www.eyeofscience.com/eos2/english/gallery/medizin/bsp5.html# Leishmania mexicana / Leishmania major] ] in the insect host, and "amastigote" (without flagella) in the vertebrate host. Infections are regarded as cutaneous, mucocutaneous, or visceral."Cutaneous" (localized and diffuse) infections appear as obvious skin reactions. The most common is the "Oriental Sore" (caused by Old World species "L. major", "L. tropica", and "L. aethiopica"). In the New World, the most common culprits are "L. mexicana" and "L. (Viannia) braziliensis". Cutaneous infections are most common in
Afghanistan ,Brazil ,Iran ,Peru ,Saudi Arabia andSyria ."Mucocutaneous" (espundia) infections will start off as a reaction at the bite, and can go via
metastasis into the mucous membrane and become fatal. Mucocutaneous infections are most common inBolivia ,Brazil andPeru . Mucocutaneous infections are also found inKaramay , China Xinjiang Uygur Autonomous Region."Visceral" infections are often recognized by fever, swelling of the liver and spleen, and
anemia . They are known by many local names, of which the most common is probably "Kala azar ", [http://homepages.uel.ac.uk/D.P.Humber/akhter/dis.htm Visceral leishmniasis: The disease] ] [http://www.bartleby.com/61/51/K0005100.html kala-azar] The American Heritage® Dictionary of the English Language ] and are caused exclusively by species of the "L. donovani" complex ("L. donovani", "L. infantum" syn. "L. chagasi"). Found in tropical and subtropical areas of all continents exceptAustralia , visceral infections are most common inBangladesh ,Brazil ,India ,Nepal andSudan . Visceral leishmaniasis also found in part of China, such as Sichuan Province, Gansu Province and Xinjiang Uygur Autonomous Region.Treatment
Antimonial compounds are the traditional treatments for leishmaniasis (
sodium stibogluconate ,meglumine antimoniate ). [ [http://www.icp.ucl.ac.be/~opperd/parasites/leish5.htm Treatment of Leishmaniasis ] ]Resistance to the antimonials is prevalent in some parts of the world, and the most common alternative is
amphotericin B [www.pdhealth.mil/downloads/Leish_brfng.ppt] (seeleishmaniasis for other treatment options). Paromomycin is an inexpensive alternative with fewer side effects than amphotericin thatThe Institute for OneWorld Health has funded for production as anorphan drug for use in treatment of leishmaniasis, starting in India.Molecular biology
An important aspect of the "Leishmania" protozoan is its
glycoconjugate layer oflipophosphoglycan (LPG). This is held together with a phosphoinositide membrane anchor, and has a tripartite structure consisting of a lipid domain, a neutral hexasaccharide, and a phosphrorylated galactose-mannose, with a termination in a neutral cap. Not only do these parasites develop post-phlebotomus digestion but, it is thought to be essential to oxidative bursts, thus allowing passage for infection. Characteristics of intracellular digestion include anendosome fusing with alysosome , releasing acidhydrolase s which degradeDNA ,RNA , proteins andcarbohydrates .Genomics
The genomes of three "Leishmania" species ("L. major", "L. infantum" and "L. braziliensis") have been sequenced, revealing more than 8300 protein-coding and 900
RNA genes. Almost 40% of protein-coding genes fall into 662 families containing between two and 500 members. Most of the smaller gene families are tandem arrays of one to three genes, while the larger gene families are often dispersed in tandem arrays at differentloci throughout thegenome . Each of the 35 or 36chromosome s are organized into a small number of gene clusters of tens-to-hundreds of genes on the same DNA strand. These clusters can be organized in head-to-head (divergent) or tail-to-tail (convergent) fashion, with the latter often separated bytRNA ,rRNA and/orsnRNA genes. Transcription of protein-coding genes initiates bi-directionally in the divergent strand-switch regions between gene clusters and extends polycistron ically through each gene cluster before terminating in the strand-switch region separating convergent clusters. "Leishmania"telomer es are usually relatively small, consisting of a few different types of repeat sequence. Evidence can be found for recombination between several different groups of telomeres. The "L. major" and "L. infantum" genomes contain only ~50 copies of inactive degenerated "Ingi"/L1Tc-related elements (DIREs), while "L. braziliensis" also contains several telomere-associated transposable elements (TATEs) and spliced leader-associated (SLACs) retroelements. The "Leishmania" genomes share a conserved core proteome of ~6200 genes with the related trypanosomatids "Trypanosoma brucei" and "Trypanosoma cruzi" , but there are ~1000 "Leishmania"-specific genes (LSGs), which are mostly randomly distributed throughout the genome. There are relatively few (~200) species-specific differences in gene content between the three sequenced "Leishmania" genomes, but ~8% of the genes appear to be evolving at different rates between the three species, indicative of different selective pressures that could be related to disease pathology. About 65% of protein-coding genes currently lack functional assignment.cite book | author = Myler P; Fasel N (editors). | title = Leishmania: After The Genome | publisher = Caister Academic Press | year = 2008 | url=http://www.horizonpress.com/leish | id = [http://www.horizonpress.com/leish ISBN 978-1-904455-28-8 ] ]Leishmania as component of CVBD
"Canine Vector-borne Diseases" (
CVBD ) covers diseases caused by pathogens transmitted by ectoparasites as ticks, fleas, sand flies or mosquitoes.Other microorganism-based diseases caused by ectoparasites include "
Bartonella ", "Borrelia ", "Babesia ", Dirofilaria, Ehrlichia, and Anaplasma.Neutrophil granulocytes - the Trojan horses for Leishmania parasites
The strategy of the "Trojan horse" as a mechanism of
pathogenicity ofintracellular microorganisms is, to avoid theimmune system and its memory function cleverly, withphagocytosis of infected andapoptotic neutrophils bymacrophages , employing the non-danger surface signals of apoptotic cells.Transmitted by the
sandfly , theprotozoan parasites of the genus "Leishmania major " may switch the strategy of the firstimmune defense from eating/inflammation/killing to eating/no inflammation/no killing of their hostphagocyte ' and corrupt it for their own benefit. They use the willingly phagocytosing polymorphonuclear neutrophil granulocytes (PMN) rigorously as a tricky hideout, where theyproliferate unrecognized from the immune system and enter the long-livedmacrophages to establish a “hidden”infection .Uptake and survival
By a
microbial infection PMN move out from the bloodstream and through the vessels’ endothelial layer, to the site of the infected tissue (dermal tissue after fly bite). They immediately start their business there as the first immune response and phagocyte the invader because of the foreign and activating surfaces. In that processes aninflammation emerges. Activated PMN secretechemokines ,IL-8 particularly, to attract furthergranulocytes and stimulate them to phagocytosis. Furthermore "Leishmania major" increases the secretion of IL-8 by PMN. In the parasites case, that may not sound reasonable at first. We can observe this mechanism on otherobligate intracellular parasites , too. For microbes like these, there are several ways to survive inside cells. Surprisingly, the co-injection of apoptotic and viable pathogens causes by far a more fulminate course of disease than injection of only viable parasites. Exposing on the surface of dead parasites the anti-inflammatory signalphosphatidylserine , usually found on apoptotic cells, "Leishmania major" switches off theoxidative burst , so killing and degradation of the co-injected viable pathogen is not achieved.In case of "Leishmania" progeny is not generated in PMN, but in this way they can survive and persist untangled on the primary site of infection. The
promastigote forms also release LCF (Leishmania chemotactic factor) to recruit actively neutrophils but not otherleukocytes , for instancemonocytes orNK cells . In addition to that, the production ofinterferon gamma (IFNγ)-inducible protein 10 (IP10) by PMN is blocked in attendance of "Leishmania", what involves the shut down of inflammatory and protective immune response by NK andTh1 cell recruitment. The pathogens stay viable during phagocytosis since their primary hosts, the PMN, expose apoptotic cell associated molecular pattern (ACAMP) signaling “no pathogen.”Persistency and attraction
The lifespan of
neutrophil granulocytes is quite short. They circulate inbloodstream for about 6 or 10 hours after leavingbone marrow , whereupon they undergo spontaneous apoptosis. Microbial pathogens have been reported to influence cellular apoptosis by different strategies. Obviously because of the inhibition ofcaspase 3-activation "Leishmania major" can induce the delay of neutrophils apoptosis and extend their lifespan for at least 2–3 days. The fact of extended lifespan is very beneficial for the development of infection because the final host cells for these parasites are macrophages, which normally migrate to the sites of infection within 2 or 3 days. The pathogens are not dronish; instead they take over the command at the primary site of infection. They induce the production by PMN of the chemokines MIP-1α and MIP-1β (macrophage inflammatory protein ) to recruit macrophages. [cite journal
quotes = no
author = Laskay T. "et al".
year = 2003
title = Neutrophil granulocytes – Trojan horses for "Leishmania major" and other intracellular microbes?
journal = Trends in Microbiology
volume = 11
issue = 5
pages = 210-4
doi = 10.1016/S0966-842X(03)00075-1
id = PMID 12781523 ]ilent phagocytosis
To save the integrity of the surrounding tissue from the
toxic cell components andproteolytic enzymes contained in neutrophils, the apoptotic PMN are silently cleared by macrophages. Dying PMN expose the "eat me"-signalphosphatidylserine which is transferred to the outer leaflet of theplasma membrane during apoptosis. By reason of delayed apoptosis the parasites that persist in PMN are taken up into macrophages, employing an absolutelyphysiological and non-phlogistic process. The strategy of this "silent phagocytosis" has following advantage for the parasite:• Taking up apoptotic cells silences macrophage killing activity leading to a survival of the pathogens.
• Pathogens inside of PMN have no direct contact to the macrophage surface
receptors , because they can not see the parasite inside the apoptotic cell. So the activation of the phagocyte for immune activation does not occur.ee Also
Canine Leishmaniasis Literature:
*Zandbergen et al. "Leishmania" disease development depends on the presence of apoptotic promastigotes in the virulent inoculum", PNAS, Sept. 2006 ( [http://www.pnas.org/cgi/reprint/103/37/13837.pdf PDF] )
*cite book | author = Shaw J. J. | title = The haemoflagellates of sloths | publisher = H. K. Lewis & Co. Ltd. | year = 1969 | url=http://fr.calameo.com/read/000009484fd0f49891ecb | id = [http://fr.calameo.com/read/000009484fd0f49891ecb ISBN 978-0718603182 ] ( [http://fr.calameo.com/read/000009484fd0f49891ecb Full text e-book] )
*cite book | author = Myler and Fasel | title = Leishmania: After The Genome | publisher = Caister Academic Press | year = 2008 | url=http://www.horizonpress.com/leish | id = [http://www.horizonpress.com/leish ISBN 978-1-904455-28-8 ]
References
External links
* The International Leishmania Network [http://www.bdt.org.br/leishnet/ (ILN)] has basic information on the disease and links to many aspects of the disease and its vector.
* A discussion list [http://www.bdt.fat.org.br/listas/leish-l/ (Leish-L)] is also available with over 600 subscribers to the list, ranging from molecular biologists to public health workers, from many countries both inside and outside endemic regions. Comments and questions are welcomed.
* [http://www.kinetoplastids.com/ KBD: Kinetoplastid Biology and Disease] , is a website devoted to leishmaniasis, sleeping sickness and
Chagas (American trypanosomiasis). It contains free access to full length peer review articles on these subjects. The site contains many articles relating to the unique kinetoplastid organelle and genetic material therein.
Wikimedia Foundation. 2010.