- Reuptake
thumb">
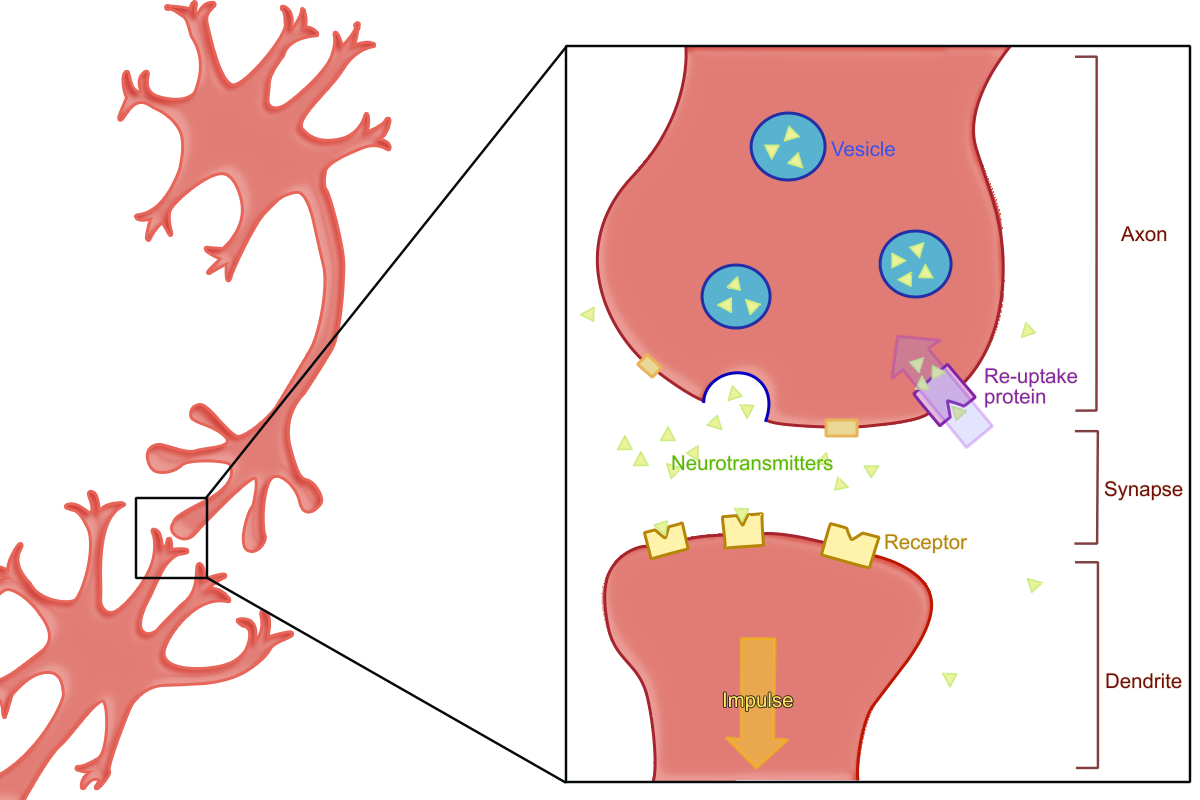
350px|A synapse during re-uptake. Note that some neurotransmitters are lost and not reabsorbed.Reuptake, or re-uptake, is the reabsorption of aneurotransmitter by theneurotransmitter transporter of a pre-synapticneuron after it has performed its function of transmitting a neural impulse.Reuptake is necessary for normal functioning because it allows for the recycling of
neurotransmitter s and regulates normal levels of neurotransmitter present in the synapse and how long a signal lasts. Because neurotransmitters are too big to diffuse through the membrane, special transport proteins are necessary for the reabsorption of neurotransmitters. Protein structure must first be determined, as it may give clues to the mechanism of reuptake.Protein Structure
Protein structure determination of a reuptake protein was not completed until roughly 1990. The technique for protein determination is contingent upon the purification, sequencing, and cloning of the transporter protein in question. After separate investigations had sequenced the DNA that coded for both GABA transporter and
norepinephrine transporter, it could be seen that there were many similarities between the two DNA sequences. Further exploration in the field of reuptake proteins found that many of the transporters associated with important neurotransmitters within the body were also very similar in sequence to the GABA and norepinephrine transporters. The members of this new family includedopamine ,norepinephrine ,serotonin , and GABA. They were given the name Classical Na+/Cl--dependent transporters.Sodium andChloride ion dependence will be discussed later in the mechanism of action. Using the commonalities among sequences and hydropathy plot analyses, it was determined that there are 12 hydrophobic membrane spanning structural units in the ‘Classical’ transporter family Masson, J., Sagné, C., Hamon, M., and Mestikawy, S.El. (1999). Neurotransmitter Transporters in the Central Nervous System. Pharmacological Reviews, 51(3), 439-464.] . In addition to this, the N and C termini exist in the intracellular space. These proteins also all have an extended fourth extracellular loop. There is an extracellular cavity in the protein, into which protrudes a helix hairpin formed by extracellular loop EL4 Zhou, Z., Zhen J., Karpowich N.K., Goetz R.M., Law C.J., Reith, M.E.A., and Wang, D. (2007). LeuT-Desipramine Structure Reveals How Antidepressants Block Neurotransmitter Reuptake. Science 317 (5843), 1390-1393.] . After using further more advanced scanning techniques, Masson et al. proposed that transmembrane unit 1 (closest to the N-terminus) exists in the membrane as a pore loop. In other words, unit 1 would not exist in a transmembrane fashion but rather as a loop existing solely in the membrane. This model has been used before to explain mechanisms for ion selectivity filters, although the purpose of this pore loop is not readily apparent. This pore loop observation is consistent across all of the classical family of transporter proteins, suggesting that it is important for some function shared among them. Later experiments indicated that atyrosine amino acid is highly conserved across transport proteins, and was shown to be essential for substrate binding and transportYamashita, A., Singh, S.K., Kawate, T., Jin, Y. & Gouaux, E. (2005) Crystal structure of a bacterial homologue of Na+/Cl--dependent neurotransmitter transporters. Nature 437, 215 – 223] . Other important features include function specific positions in the transmembrane section 1, where anaspartate differentiates betweenmonoamine substrates and aglycine differentiates betweenamino acid substrates. It has also been shown that a negatively charged residue in the extracellular loop 5 and the transmembrane domain 10 may form an external gate. Yamashita et al. chose to examine a bacterial homologue (LeuT: Leucine transporter) of these classical transporters from the bacterium "Aquifex aeolicus ". Protein structure of LeuT was determined via crystallization. They chose to crystallize the system withleucine and its respective 2 sodium ions in order to visualize the pore and binding sites. The crystal was scanned viamulti-wavelength anomalous dispersion . Using this technique, they were able to replicate the findings in the classical transporters, but they determined that the folding and final structure were unique. They found that the transmembrane domains 1 and 6 contain unwound segments in the middle of the membrane. Along with this, transmembrane domains 3 and 8 and the areas surrounding the unwound sections of 1 and 6 form the substrate and sodium ion binding sites. They observed pseudo-symmetry among the LeuT protein, seen best when observing transmembrane domains 1-5 against 6-10.Mechanism of Action
The classic transporter proteins utilize
symport to transport neurotransmitter across the membrane of the presynaptic neuron. The classical transporters, which are sodium and chloride ion dependent, take advantage of the large sodium gradients across the membrane. The neurotransmitter in question will bind to sodium ions, where the sodium ion will flow down its concentration gradient as well as electrical gradient. These forces will pull the neurotransmitter into the cell, against its own gradients. The chloride ion also contributes by flowing down its concentration gradient, but it flows against the electrical gradient, greatly reducing its efficiency as a symport ion. The role of the chloride ion in the symport mechanism is not exactly known, but has been implicated to be useful for counterbalancing the charge of the sodium symport ionsZomot, E., Bendahan, A., Quick, M., Zhao, Y., Javitch, J.A. & Kanner, B.I. (2007). Mechanism of chloride interaction with neurotransmitter:sodium symporters. Nature, 449, 726-730.] . Because the neurotransmitter binds to sodium ions, their respective binding sites are very close in proximity. In the LeuT protein, the binding sites would be across transmembrane domains 3 and 8, and specifically the unwound sections of 1 and 6. These binding sites are made up of hydrogen and ionic bonding between the substrate and the transport protein. The primary sodium binding sites are Na1 and Na2, which exist in the transmembrane domains 1 (TM1) and 6 (TM6) in the unwound regions. In the LeuT example, 2 sodium ions bind to the substrate leucine. These 2 sodium ions work to pull leucine into the cell, but also by stabilizing the core of LeuT. Sodium ion 1 (binds to Na1) is octahedrally coordinated by the leucine carboxy oxygen, the carbonyl oxygens of Ala22 (transmembrane domain 1), Thr254 (TM6), side-chain carbonyl oxygens of Asn27 (TM1) and Asn286 (TM7), and the hydroxyl oxygen of Thr254 (TM6). The second sodium ion (binds to Na2) is trigonal bi-pyramidally coordinated by means of the carbonyl oxygens of Gly20 and Val23 (TM1), Ala351 (TM8), and the hydroxyl oxygens from Thr354 and Ser355 (TM8).After sodium ion coordination has taken place, some conformational change must occur. In an isolated system of TM3, TM8, TM1, and TM6, the unwound sections of 1 and 6 act like a joint and pivot the entire transmembrane domain relative to the rest of the protein and TM3 and TM8. The summation of actions mimics a half revolving door. Both TM3 and TM8 remain stationary and TM1 and TM6 swing about it.
The resulting system has affinity for both the substrate and sodium ions at both the intracellular level and the extracellular level. As the system progresses, the extracellular openings are blocked off by a gate. Because of the sodium gradient, it is unlikely that much substrate transfer occurs from the intracellular level to the extracellular level. The LeuT transporter protein is a homologue of the human classical transport proteins, and thus the proposed mechanism of action can be assumed to be largely similar to the human model.
Mechanism of Reuptake Inhibition
The main objective of a
reuptake inhibitor is to substantially decrease the rate by which neurotransmitters are reabsorbed into the presynaptic neuron, leaving a net gain in the concentration of neurotransmitter in the synapse. This increases the probability and frequency of neurotransmitter binding to postsynaptic neurotransmitter receptors. Depending on the neurological system in question, a reuptake inhibitor can have drastic effects on cognition and behavior. Zhou et al. examined howtricyclic antidepressant s act upon a reuptake protein in order to inhibit reuptake of the appropriate neurotransmitters. They chose to examine the LeuT system as it was previously crystallized by Yamashita et al. in 2005. They also chose to examine LeuT because it is a homologue of the transport proteins of concern when considering depression. They examined this relationship between tricyclic antidepressants and the LeuT system by crystallizing the two together, along with leucine and the respective 2 sodium ions. They found that the leucine and the sodium ions bound exactly as previously determined by Yamashita et al. They also observed that the tricyclic antidepressant (desipramine ) followed leucine into the pore. Desipramine effectively sits on top of leucine in LeuT and they both bind to the same protein residue. The tail of desipramine extends into the extracellular space. Most of the LeuT structure does not change, but the EL4 hairpin does form around desipramine as if it were locking it in place in the LeuT pore. Desipramine also acts by binding to the extracellular gate and forming a salt bridge between TM1 and TM10. This action prevents TM1 from pivoting, which prevents leucine from ever being exposed to the intracellular side of LeuT. Desipramine works as a reuptake inhibitor by effectively blocking and crowding up the pore of LeuT, which physically prevents its normal course of action.Human Systems
Horschitz et al.Horschitz, S., Hummerich, R., Schloss, P. (2001). Structure, function and regulation of the 5-hydroxytryptamine (serotonin) transporter. Biochemical Society Transactions, 29(6), 728-732.] examined reuptake inhibitor selectivity among the rat serotonin reuptake protein (SERT) expressed in human embryonic kidney cells (HEK-SERT). They presented SERT with varying doses of either citalopram (SSRI) or desipramine (an inhibitor of norepinephrine reuptake protein, NET). By examining the dose-response curves (using a normal medium as control), they were able to quantify that
citalopram acted on SERT as a SSRI, and that desipramine had no effect on SERT. In a separate experiment, Horschitz et al.. exposed HEK-SERT with citalopram on a long-term basis. They noticed that long-term exposure led to a down-regulation of binding sites. These results suggest some mechanism for long-term changes in the pre-synaptic neuron after drug therapy. Horschitz et al. found that after removing citalopram from the system, normal levels of SERT binding site expression returned. Depression has been suggested to be a result of a decrease of serotonin found in the synapse. This theory has been supported by the successful reduction of depressive symptoms after administration of tri-cyclic antidepressants (such as desipramine) and SSRI’s. Tri-cyclic antidepressants inhibit the reuptake of both serotonin and norepinephrine by acting upon both the SERT and NET. SSRI’s selectively inhibit the reuptake of serotonin by acting upon SERT. The net result is an increased amount of serotonin in the synapse, thus increasing the probability that serotonin will interact with a serotonin receptor of the postsynaptic neuron. There are additional mechanisms by which serotonin autoreceptor desensitization must occur, but the net result is the same [Członkowska, A.I., Zienowicz, M., Bidziński, A., Maciejak, P., Lehner, M, Taracha, E., Wisłowska, A., Płaźnik, A.. (2003). The role of neurosteroids in the anxiolytic, antidepressive- and anticonvulsive effects of selective serotonin reuptake inhibitors. Med Sci Monit; 9(11): 270-275] . This increases serotonin signaling, which then acts to elevate mood and thus relieve depressive symptoms. The net effect ofamphetamine (AMPH) use is an increase of dopamine in the synapse. It has been shown that AMPH acts upon the dopamine reuptake protein (DAT) in a reverse fashion. In DAT knockout mice, dopamine levels in the synapse (as measured by microdialysis) were no different when they were exposed to AMPH relative to baseline levelsJones, S.R., Gainetdinov, R.R., Wightman R.M., and Caron, M.G. (1998). Mechanisms of Amphetamine Action Revealed in Mice Lacking the Dopamine Transporter. The Journal of Neuroscience, 18(6):1979–1986] . In normal mice, levels of dopamine in the synapse rose to ten times normal levels after exposure to AMPH.Neuroprotective Role
Astrocytes seem to utilize reuptake mechanisms for a neuroprotective role. Astrocytes use GLT-1 to removeglutamate from the synapse. GLT-1 knockout mice were more prone to lethal and spontaneous seizures and acute brain injuries among the cortex. These effects could be linked to increased concentrations of glutamate in the brains of GLT-1 knockout mice, analyzed post-mortem [Tanaka, K., Watase, K., Manabe, T., Yamada, K., Watanabe, M., Takahashi, K., Iwama, H., Nishikawa, T, Ichihara, N., Kikuchi,T., Okuyama, S., Kawashima, N., Hori, S., Takimoto, M., Wada, K. (1997). Epilepsy and Exacerbation of Brain Injury in Mice Lacking the Glutamate Transporter GLT-1. Science 276, 1699-1702.] .References
Wikimedia Foundation. 2010.