- Dengue virus
-
Dengue virus 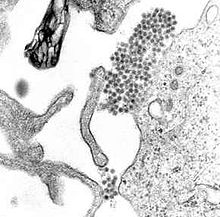
A TEM micrograph showing Dengue virus virions (the cluster of dark dots near the center). Virus classification Group: Group IV ((+)ssRNA) Family: Flaviviridae Genus: Flavivirus Species: Dengue virus Dengue virus (DENV) in one of four serotypes is the cause of dengue fever. It is a mosquito-borne single positive-stranded RNA virus of the family Flaviviridae; genus Flavivirus.[1][2] All four serotypes can cause the full spectrum of disease.[1]
Its genome is about 11000 bases that codes for three structural proteins, capsid protein C, membrane protein M, envelope protein E; seven nonstructural proteins, NS1, NS2a, NS2b, NS3, NS4a, NS4b, NS5; and short non-coding regions on both the 5' and 3' ends.[1][3] Further classification of each serotype into genotypes often relates to the region where particular strains are commonly found or were first found.
Contents
Evolution
The Dengue 1 virus appears to have evolved in the early 19th century[4] Based on the analysis of the envelope protein there are at least four genotypes (1 to 4). The rate of nucleotide substitution for this virus has been to be 6.5×10-4 per nucleotide per year, a rate similar to other RNA viruses. The American African genotype has been estimated to have evolved between 1907 to 1949. This period includes World War I and II which were associated with considerable movement of populations and environmental disturbance, factors known to promote the evolution of new vector borne viral species.
Life cycle
The primary life cycle of the dengue virus involves humans and mosquitoes.[5] There is also another cycle in Africa and also on Asia, which involves non human primates.[5]
Structure
E protein
The DENV E (envelope) protein, found on the viral surface, is important in the initial attachment of the viral particle to the host cell. Several molecules which interact with the viral E protein (ICAM3-grabbing non-integrin.,[6] CD209 ,[7] Rab 5 ,[8] GRP 78 ,[9] and The Mannose Receptor [10])have been shown to be important factors mediating attachment and viral entry.[11]
prM/M protein
The DENV prM (membrane) protein, which is important in the formation and maturation of the viral particle, consists of seven antiparallel β-strands stabilized by three disulphide bonds.[11]
The glycoprotein shell of the mature DENV virion consists of 180 copies each of the E protein and M protein. The immature virion starts out with the E and prM proteins forming 90 heterodimers that give a spiky exterior to the viral particle. This immature viral particle buds into the endoplasmic reticulum and eventually travels via the secretory pathway to the Golgi apparatus. As the virion passes through the trans-Golgi Network (TGN) it is exposed to low pH. This acidic environment causes a conformational change in the E protein which disassociates it from the prM protein and causes it to form E homodimers. These homodimers lie flat against the viral surface giving the maturing virion a smooth appearance. During this maturation pr peptide is cleaved from the M peptide by the host protease, furin. The M protein then acts as a transmembrane protein under the E-protein shell of the mature virion. The pr peptide stays associated with the E protein until the viral particle is released into the extracellular environment. This pr peptide acts like a cap, covering the hydrophobic fusion loop of the E protein until the viral particle has exited the cell.[11]
NS3 protein
The DENV NS3 is a serine protease, as well as an RNA helicase and RTPase/NTPase. The protease domain consists of six β-strands arranged into two β-barrels formed by residues 1-180 of the protein. The catalytic triad (His-51, Asp-75 and Ser-135), is found between these two β-barrels, and its activity is dependent on the presence of the NS2B cofactor. This cofactor wraps around the NS3 protease domain and becomes part of the active site. The remaining NS3 residues (180-618), form the three subdomains of the DENV helicase. A six-stranded parallel β-sheet surrounded by four α-helices make up subdomains I and II, and subdomain III is composed of 4 α-helices surrounded by three shorter α-helices and two antiparallel β-strands.[11]
NS5 protein
The DENV NS5 protein is a 900 residue peptide with a methyltransferase domain at its N-terminal end (residues 1-296) and a RNA-dependent RNA polymerase (RdRp) at its C-terminal end (residues 320–900). The methyltransferase domain consists of an α/β/β sandwich flanked by N-and C-terminal subdomains. The DENV RdRp is similar to other RdRps containing palm,, finger, and thumb subdomains and a GDD motif for incorporating nucleotides.[11]
Severe disease
The reason that some people suffer from more severe forms of dengue, such as dengue hemorrhagic fever, is multifactorial. Different strains of viruses interacting with people with different immune backgrounds lead to a complex interaction. Among the possible causes are cross-serotypic immune response, through a mechanism known as antibody-dependent enhancement, which happens when a person who has been previously infected with dengue gets infected for the second, third or fourth time. The previous antibodies to the old strain of dengue virus now interfere with the immune response to the current strain, leading paradoxically to more virus entry and uptake.[12]
References
- ^ a b c Rodenhuis-Zybert IA, Wilschut J, Smit JM (August 2010). "Dengue virus life cycle: viral and host factors modulating infectivity". Cell. Mol. Life Sci. 67 (16): 2773–86. doi:10.1007/s00018-010-0357-z. PMID 20372965.
- ^ WHO (2009). Dengue Guidelines for Diagnosis, Treatment, Prevention and Control. World Health Organization. ISBN 9241547871. http://whqlibdoc.who.int/publications/2009/9789241547871_eng.pdf.
- ^ Hanley, K.A. and Weaver, S.C. (editors) (2010). Frontiers in Dengue Virus Research. Caister Academic Press. ISBN 978-1-904455-50-9.
- ^ Patil JA, Cherian S, Walimbe AM, Patil BR, Sathe PS, Shah PS, Cecilia D (2011) Evolutionary dynamics of the American African genotype of dengue type 1 virus in India (1962-2005). Infect Genet Evol
- ^ a b "Dengue virus". http://ci.vbi.vt.edu/pathinfo/pathogens/Dengue1.html.
- ^ Tassaneetrithep B, Burgess TH, Granelli-Piperno A, et al. (April 2003). "DC-SIGN (CD209) mediates dengue virus infection of human dendritic cells". J. Exp. Med. 197 (7): 823–9. doi:10.1084/jem.20021840. PMC 2193896. PMID 12682107. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2193896.
- ^ Krishnan MN, Sukumaran B, Pal U, et al. (May 2007). "Rab 5 is required for the cellular entry of dengue and West Nile viruses". J. Virol. 81 (9): 4881–5. doi:10.1128/JVI.02210-06. PMC 1900185. PMID 17301152. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1900185.
- ^ Jindadamrongwech S, Thepparit C, Smith DR (May 2004). "Identification of GRP 78 (BiP) as a liver cell expressed receptor element for dengue virus serotype 2". Arch. Virol. 149 (5): 915–27. doi:10.1007/s00705-003-0263-x. PMID 15098107.
- ^ Miller JL, de Wet BJ, deWet BJ, et al. (February 2008). "The mannose receptor mediates dengue virus infection of macrophages". PLoS Pathog. 4 (2): e17. doi:10.1371/journal.ppat.0040017. PMC 2233670. PMID 18266465. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2233670.
- ^ a b c d e Perera R, Kuhn RJ (August 2008). "Structural proteomics of dengue virus". Curr. Opin. Microbiol. 11 (4): 369–77. doi:10.1016/j.mib.2008.06.004. PMC 2581888. PMID 18644250. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2581888.
- ^ Dejnirattisai W, Jumnainsong A, Onsirisakul N, et al. (May 2010). "Cross-reacting antibodies enhance dengue virus infection in humans". Science 328 (5979): 745–8. Bibcode 2010Sci...328..745D. doi:10.1126/science.1185181. PMID 20448183.
Categories:- Flaviviruses
Wikimedia Foundation. 2010.