- Collective animal behavior
-
 Sort sol. Starling flock at sunset in Denmark
Sort sol. Starling flock at sunset in Denmark
Collective animal behavior describes the coordinated behavior of large groups of similar animals and the emergent properties of these groups. Facets of this topic include the costs and benefits of group membership, the transfer of information across the group, the group decision-making process, and group locomotion and synchronization. Studying the principles of collective animal behavior has relevance to human engineering problems through the philosophy of biomimetics. For instance, determining the rules by which an individual animal navigates relative to its neighbors in a group can lead to advances in the deployment and control of groups of swimming or flying micro-robots such as UAVs (Unmanned Aerial Vehicles).
Contents
Examples
Examples of collective animal behavior include:
- Flocking birds
- Herding ungulates
- Shoaling and schooling fish
- Swarming Antarctic krill
- Pods of dolphins
- Marching locusts
- Nest building ants
Proposed functions
Many functions of animal aggregations have been proposed. These proposed functions may be grouped into the four following categories: social and genetic, anti-predator, enhanced foraging, and increased locomotion efficiency.
Social interaction
Support for the social and genetic function of aggregations, especially those formed by fish, can be seen in several aspects of their behavior. For instance, experiments have shown that individual fish removed from a school will have a higher respiratory rate than those found in the school. This effect has been attributed to stress, and the effect of being with conspecifics therefore appears to be a calming one and a powerful social motivation for remaining in an aggregation.[1] Herring, for instance, will become very agitated if they are isolated from conspecifics.[2] Fish schools have also been proposed to serve a reproductive function since they provide increased access to potential mates.
Protection from predators
 School of goldband fusiliers
School of goldband fusiliersSeveral anti-predator functions of animal aggregations have been proposed. One potential method by which fish schools or bird flocks may thwart predators is the ‘predator confusion effect’ proposed and demonstrated by Milinksi and Heller (1978).[3] This theory is based on the idea that it becomes difficult for predators to pick out individual prey from groups because the many moving targets create a sensory overload of the predator's visual channel.
A second potential anti-predator effect of animal aggregations is the ‘many eyes’ hypothesis. This theory states that as the size of the group increases, the task of scanning the environment for predators can be spread out over many individuals. Not only does this mass collaboration presumably provide a higher level of vigilance, it could also allow more time for individual feeding.[4][5]
A third hypothesis for an anti-predatory effect of animal aggregation is the ‘encounter dilution’ effect. Hamilton, for instance, proposed that the aggregation of animals was due to a “selfish” avoidance of a predator and was thus a form of cover-seeking.[6] Another formulation of the theory was given by Turner and Pitcher and was viewed as a combination of detection and attack probabilities.[7] In the detection component of the theory, it was suggested that potential prey might benefit by living together since a predator is less likely to chance upon a single group than a scattered distribution. In the attack component, it was thought that an attacking predator is less likely to eat a particular animal when a greater number of individuals are present. In sum, an individual has an advantage if it is in the larger of two groups, assuming that the probability of detection and attack does not increase disproportionately with the size of the group.[8]
Enhanced foraging
A third proposed benefit of animal groups is that of enhanced foraging. This ability was demonstrated by Pitcher and others in their study of foraging behavior in shoaling cyprinids.[9] In this study, the time it took for groups of minnows and goldfish to find a patch of food was quantified. The number of fishes in the groups was varied, and a statistically significant decrease in the amount of time necessary for larger groups to find food was established. Further support for an enhanced foraging capability of schools is seen in the structure of schools of predatory fish. Partridge and others analyzed the school structure of Atlantic bluefin tuna from aerial photographs and found that the school assumed a parabolic shape, a fact that was suggestive of cooperative hunting in this species (Partridge et al., 1983).[10]
Increased locomotion efficiency
This theory states that groups of animals moving in a fluid environment may save energy when swimming or flying together, much in the way that bicyclists may draft one another in a peloton. Geese flying in a Vee formation are also thought to save energy by flying in the updraft of the wingtip vortex generated by the previous animal in the formation. Ducklings have also been shown to save energy by swimming in a line.[11] Increased efficiencies in swimming in groups have also been proposed for schools of fish and Antarctic krill.
Group structure
The structure of large animal groups has been difficult to study because of the large number of animals involved. The experimental approach is therefore often complemented by mathematical modeling of animal aggregations.
Experimental approach
Experiments investigating the structure of animal aggregations seek to determine the 3D position of each animal within a volume at each point in time. It is important to know the internal structure of the group because that structure can be related to the proposed motivations for animal grouping. This capability requires the use of multiple cameras trained on the same volume in space, a technique known as stereophotogrammetry. When hundreds or thousands of animals occupy the study volume, it becomes difficult to identify each one. In addition, animals may block one another in the camera views, a problem known as occlusion. Once the location of each animal at each point in time is known, various parameters describing the animal group can be extracted.
These parameters include:
Density: The density of an animal aggregation is the number of animals divided by the volume (or area) occupied by the aggregation. Density may not be a constant throughout the group. For instance, starling flocks have been shown to maintain higher densities on the edges than in the middle of the flock, a feature that is presumably related to defense from predators.[12]
Polarity: The group polarity describes if the group animals are all pointing in the same direction or not. In order to determine this parameter, the average orientation of all animals in the group is determined. For each animal, the angular difference between its orientation and the group orientation is then found. The group polarity is then the average of these differences (Viscido 2004).[13]
Nearest Neighbor Distance: The nearest neighbor distance (NND) describes the distance between the centroid of one animal (the focal animal) and the centroid of the animal nearest to the focal animal. This parameter can be found for each animal in an aggregation and then averaged. Care must be taken to account for the animals located at the edge of an animal aggregation. These animals have no neighbor in one direction.
Nearest Neighbor Position: In a polar coordinate system, the nearest neighbor position describes the angle and distance of the nearest neighbor to a focal animal.
Packing Fraction: Packing fraction is a parameter borrowed from physics to define the organization (or state i.e. solid, liquid, or gas) of 3D animal groups. It is an alternative measure to density. In this parameter, the aggregation is idealized as an ensemble of solid spheres, with each animal at the center of a sphere. The packing fraction is defined as the ratio of the total volume occupied by all individual spheres divided by the global volume of the aggregation (Cavagna 2008). Values range from zero to one, where a small packing fraction represents a dilute system like a gas. Cavagna found that the packing fraction for groups of starlings was 0.012.[14]
Integrated Conditional Density: This parameter measures the density at various length scales and therefore describes the homogeneity of density throughout an animal group.[14]
Pair Distribution Function: This parameter is usually used in physics to characterize the degree of spatial order in a system of particles. It also describes the density, but this measures describes the density at a distance away from a given point. Cavagna et al. found that flocks of starlings exhibited more structure than a gas but less than a liquid.[14]
Modeling approach
The simplest mathematical models of animal aggregations generally instruct the individual animals to follow three rules:
- Move in the same direction as your neighbor
- Remain close to your neighbors
- Avoid collisions with your neighbors
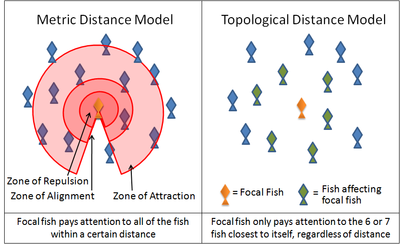 A diagram illustrating the difference between 'metric distance' and 'topological distance' in reference to fish schools
A diagram illustrating the difference between 'metric distance' and 'topological distance' in reference to fish schoolsAn example of such a simulation is the Boids program created by Craig Reynolds in 1986. Another is the Self Propelled Particle model. Many current models use variations on these rules. For instance, many models implement these three rules through layered zones around each animal. In the zone of repulsion very close to the animal, the focal animal will seek to distance itself from its neighbors in order to avoid a collision. In the slightly further away zone of alignment, a focal animal will seek to align its direction of motion with its neighbors. In the outmost zone of attraction, which extends as far away from the focal animal as it is able to sense, the focal animal will seeks to move towards a neighbor. The shape of these zones will necessarily be affected by the sensory capabilities of the animal. For example, the visual field of a bird does not extend behind its body. Fish, on the other hand, rely on both vision and on hydrodynamic signals relayed through its lateral line. Antarctic krill rely on vision and on hydrodynamic signals relayed through its antennae.
Recent studies of starling flocks have shown, however, that each bird modifies its position relative to the six or seven animals directly surrounding it, no matter how close or how far away those animals are.[15] Interactions between flocking starlings are thus based on a topological rule rather than a metric rule. It remains to be seen whether the same rule can be applied to other animals.
Collective decision making
Aggregations of animals are faced with decisions which they must make if they are to remain together. For a school of fish, an example of a typical decision might be which direction to swim when confronted by a predator. Social insects such as ants and bees must collectively decide where to build a new nest.[16] A herd of elephants must decide when and where to migrate. How are these decisions made? Do stronger or more experienced 'leaders' exert more influence than other group members, or does the group make a decision by consensus? The answer probably depends on the species. While the role of a leading matriarch in an elephant herd is well known, studies have shown that some animal species use a consensus approach in their collective decision-making process.
A recent investigation showed that small groups of fish used consensus decision-making when deciding which fish model to follow. The fish did this by a simple quorum rule such that individuals watched the decisions of others before making their own decisions. This technique generally resulted in the 'correct' decision but occasionally cascaded into the 'incorrect' decision. In addition, as the group size increased, the fish made more accurate decisions in following the more attractive fish model.[17] Consensus decision-making, a form of collective intelligence, thus effectively uses information from multiple sources to generally reach the correct conclusion.
Some simulations of collective decision-making use the Condorcet method to model the way groups of animals come to consensus.
See also
- Biomimetics
- Collective intelligence
- Swarm intelligence
- Emergence
References
- ^ Abrahams, M. and Colgan, P. Risk of predation, hydrodynamic efficiency, and their influence on school structure. Environmental Biol of Fishes 13, 3, pp 195-202, 1985.
- ^ Partridge, B., Pitcher, T., Cullen, M., and Wilson, J. The three-dimensional structure of fish schools. Behav Ecol and Sociobiology 6:4, pp 277-288. 1980.
- ^ Milinski, H. and Heller, R. Influence of a predator on the optimal foraging behavior of sticklebacks. Nature 275, pp642-644. 1978.
- ^ Roberts, G. Why individual vigilance increases as group size increases. Anim Behav. 51. pp 1077-1086. 1996.
- ^ Lima, S. Back to the basics of anti-predatory vigilance: the group-size effect. Animal Behaviour 49:1. pp 11-20. 1995.
- ^ Hamilton, W. D. Geometry for the selfish herd. J. Theor Biology 31, pp 295-311. 1971
- ^ Turner, G. and Pitcher, T. Attack abatement: a model for group protection by combined avoidance and dilution. American Naturalist 128:2, pp228-240. 1986.
- ^ Krause, J., Ruxton, G., and Rubenstein, D. Is there always an influence of shoal size on predator hunting success? Journal of Fish Biology 52, pp 494-501. 1998.
- ^ Pitcher, T., Magurran, A., and Winfield, I. Fish in larger shoals find food faster. Behav. Ecol. and Sociobiology 10:2, 1982.
- ^ Partridge, B. Johansson, J and Kalish, J. The structure of schools of giant bluefin tuna in Cape Cod Bay. Environmental Biology of Fishes 9: 3-4. pp 253. 1983.
- ^ Fish, F.E. Kinematics of ducklings swimming in formation: consequences of position. (1995) Journal of Experimental Zoology 273:1-11.
- ^ M. Ballerini, N. Cabibbo, R. Candelier, A. Cavagna, E. Cisbani, I. Giardina, A. Orlandi, G. Parisi, A. Procaccini, M. Viale, and V. Zdravkovic (2008) ‘Empirical investigation of starling flocks: a benchmark study in collective animal behaviour’ Animal Behavior 76(1): 201-215.
- ^ Viscido, S., Parrish, J., and Grunbaum, D. (2004) Individual behavior and emergent properties of fish schools: a comparison of observation and theory. Marine Ecology Progress Series 273:239-249.
- ^ a b c Cavagna, A., Cimarelli, Giardina, Orlandi, Parisi, Procaccini, Santagati and Stefanini. (2008). New statistical tools for analyzing the structure of animal groups. Mathematical Biosciences 214:32-37.
- ^ M. Ballerini, N. Cabibbo, R. Candelier, A. Cavagna, E. Cisbani, I. Giardina, V. Lecomte, A. Orlandi, G. Parisi, A. Procaccini, M. Viale, and V. Zdravkovic (2008) ‘Interaction ruling animal collective behavior depends on topological rather than metric distance: Evidence from a field study’ PNAS 105:1232-1237.
- ^ Sumpter, D. http://www.collective-behavior.com/index.html
- ^ Sumpter, D., Krause, J,. James, R., Couzin, I., and Ward, A. (2008). Consensus decision making by fish. Current Biology 18:1773-1777.
Recommended reading
Camazine, S., Deneubourg, J.L., Franks, N.R., Sneyd, J., Theraulaz, G. and Bonabeau, E. (2001) Self-Organization in Biological Systems Princeton University Press, Princeton, N.J. ISBN 0691012113 (especially Chapter 11)
Sumpter, D. J. T. (2010) "Collective Animal Behavior" Princeton University Press, Pinceton, ISBN13: 978-0-691-14843-4
External links
- Collective Animal Behavior website organized around David Sumpter's book (2008) by the same name
- STARFLAG project: Description of starling flocking project
- Center for Biologically Inspired Design at Georgia Tech
- David Sumpter's research website
- Iain Couzin's research website
- Website of Julia Parrish, an animal aggregation researcher
- Research for this Wikipedia entry was conducted as a part of a Locomotion Neuromechanics course (APPH 6232) offered in the School of Applied Physiology at Georgia Tech
Swarming Swarm algorithms - Agent-based models
- Ant colony optimization
- Ant robotics
- Artificial Ants
- Bees algorithm
- Bee colony optimization
- Boids
- Crowd simulation
- Firefly algorithm
- Glowworm swarm optimization
- Particle swarm optimization
- Self-propelled particles
- Swarm intelligence
- Swarm (simulation)

Biological swarming - Agent-based model in biology
- Bait ball
- Collective animal behavior
- Feeding frenzy
- Flock
- Flocking
- Herd
- Herd behavior
- Mixed-species foraging flock
- Mobbing behavior
- Pack hunter
- Patterns of self-organization in ants
- Sardine run
- Shoaling and schooling
- Sort sol
- Swarming behaviour
- Swarming (honey bee)
- Swarming motility
Animal migration Swarm robotics Related topics Categories:



Wikimedia Foundation. 2010.