- Morpheein
-
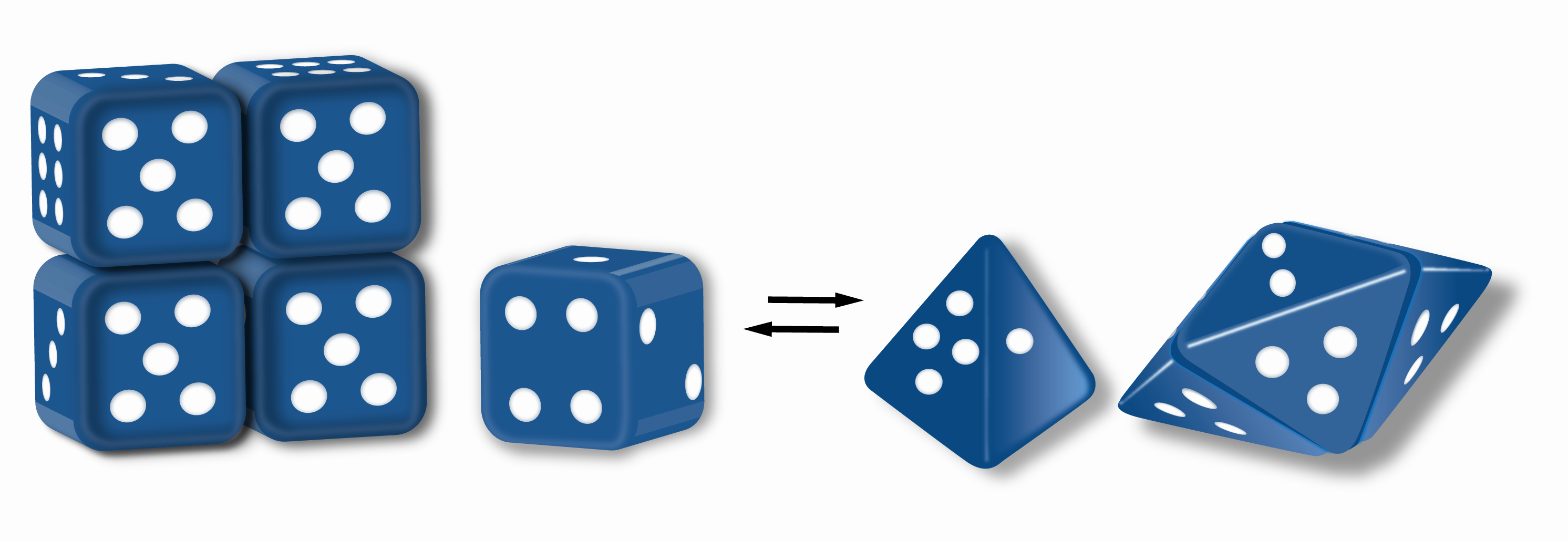
 Proteins that function as morpheeins are illustrated using a dice analogy where one die can morph to two different shapes, cubic and tetrahedral. The illustrated assemblies apply a rule that the die face with one spot must contact the die face with four spots. To satisfy the rule for each die in an assembly, the cubic die can only form a tetramer and the tetrahedral die can only assemble to a pentamer. This is analogous to two different conformations (morpheein forms) of a protein subunit each dictating assembly to a different oligomer. All dice in one assembly must be of the same shape before assembly. Thus, for example, the tetramer must come apart, and its component dice must change shape to a pyramid before they can participate in assembly into a pentamer.
Proteins that function as morpheeins are illustrated using a dice analogy where one die can morph to two different shapes, cubic and tetrahedral. The illustrated assemblies apply a rule that the die face with one spot must contact the die face with four spots. To satisfy the rule for each die in an assembly, the cubic die can only form a tetramer and the tetrahedral die can only assemble to a pentamer. This is analogous to two different conformations (morpheein forms) of a protein subunit each dictating assembly to a different oligomer. All dice in one assembly must be of the same shape before assembly. Thus, for example, the tetramer must come apart, and its component dice must change shape to a pyramid before they can participate in assembly into a pentamer.
Morpheein – a protein that can form two or more different homo-oligomers (morpheein forms), but must come apart and change shape to convert between forms. The alternate shape may reassemble to a different oligomer. The shape of the subunit dictates which oligomer is formed.[1] The conformational changes necessary to dictate oligomer formation need not be very extensive.[1][2] Each oligomer has a finite number of subunits (stoichiometry). Morpheeins can interconvert between forms under physiological conditions and can exist as an equilibrium of different oligomers. These oligomers are physiologically relevant and are not misfolded proteins; this distinguishes morpheeins from prions. The different oligomers have distinct functionality. Interconversion of morpheein forms can be a structural basis for allosteric regulation.[1][2][3] A mutation that shifts the normal equilibrium of morpheein forms can serve as the basis for a conformational disease.[4] Features of morpheeins can be exploited for drug discovery.[1][3] The dice image (Fig 1) represents a morpheein equilibrium containing two different monomeric shapes that dictate assembly to a tetramer or a pentamer. The one protein that is established to function as a morpheein is porphobilinogen synthase,[2][5] though there are suggestions throughout the literature that other proteins may function as morpheeins (for more information please see "Morpheeins Database" in #External links).
Contents
Implications for drug discovery
Conformational differences between subunits of different oligomers and related functional differences of a morpheein provide a starting point for drug discovery. Protein function is dependent on the oligomeric form; therefore, the protein’s function can be regulated by shifting the equilibrium of forms. A small molecule compound can shift the equilibrium either by blocking or favoring formation of one of the oligomers. The equilibrium can be shifted using a small molecule that has a preferential binding affinity for only one of the alternate morpheein forms. An inhibitor of porphobilinogen synthase with this mechanism of action has been documented.[3]
Implications for allosteric regulation
The morpheein model of allosteric regulation has similarities to and differences from other models.[1][6] The concerted model (the Monod, Wyman and Changeux (MWC) model) of allosteric regulation requires all subunits to be in the same conformation or state within an oligomer like the morpheein model.[7][8] However, neither this model nor the sequential model (Koshland, Nemethy, and Filmer model) takes into account that the protein must dissociate to interconvert between oligomers.[7][8][9][10]
Implications for teaching about protein structure-function relationships
It is generally taught that a given amino acid sequence will have only one physiologically relevant (native) quaternary structure; morpheeins challenge this concept. The morpheein model does not require gross changes in the basic protein fold.[1] The conformational differences that accompany conversion between oligomers may be similar to the protein motions necessary for function of some proteins.[11] Graduate, undergraduate and possibly even high school biochemistry students could benefit from presentation of morpheeins. The morpheein model highlights the importance of conformational flexibility for protein functionality and offers a potential explanation for proteins showing non-Michaelis-Menten kinetics, hysteresis, and/or protein concentration dependence to its specific activity.[6]
Implications for understanding the structural basis for disease
The term “conformational disease” generally encompasses only mutations that result in misfolded proteins that aggregate, such as Alzheimer’s and Creutzfeldt-Jakob diseases.[12] In light of the discovery of morpheeins, however, this definition could be expanded to include mutations that shift an equilibrium of alternate oligomeric forms of a protein. An example of such a conformational disease is ALAD porphyria, which results from a mutation of porphobilinogen synthase that causes a shift in its morpheein equilibrium.[4]
References
- ^ a b c d e f Jaffe, E. K. (2005). "Morpheeins - a new structural paradigm for allosteric regulation." Trends Biochem. Sci. 30(9): 490-497.
- ^ a b c Breinig, S., J. Kervinen, et al. (2003). "Control of tetrapyrrole biosynthesis by alternate quaternary forms of porphobilinogen synthase." Nat. Struct. Biol. 10(9): 757-763.
- ^ a b c Lawrence, S. H., U. D. Ramirez, et al. (2008). "Shape shifting leads to small-molecule allosteric drug discovery." Chem. Biol. 15(6): 586-596.
- ^ a b Jaffe, E. K. and L. Stith (2007). "ALAD Porphyria Is a Conformational Disease." Am J Hum Genet 80(2): 329-337.
- ^ Tang, L., L. Stith, et al. (2005). "Substrate-induced interconversion of protein quaternary structure isoforms." J. Biol. Chem. 280(16): 15786-15793.
- ^ a b Lawrence, S. H. and E. K. Jaffe (2008). "Expanding the concepts in protein structure-function relationships and enzyme kinetics: Teaching using morpheeins." Biochemistry and Molecular Biology Education 36(4): 274-283.
- ^ a b Monod, J., J. P. Changeux, et al. (1963). "Allosteric proteins and cellular control systems." J Mol Biol 6: 306-29.
- ^ a b Monod, J., J. Wyman, et al. (1965). "On the nature of allosteric transitions: A plausible model." J. Mol. Biol. 12: 88-118.
- ^ Koshland, D. E., Jr., G. Nemethy, et al. (1966). "Comparison of experimental binding data and theoretical models in proteins containing subunits." Biochemistry 5(1): 365-85.
- ^ Koshland, D. E., Jr (1970). The molecular basis for enzyme regulation. The enzymes. P. D. Boyer. New York, Academic Press. 1: 341-396.
- ^ Gerstein, M. and N. Echols (2004). "Exploring the range of protein flexibility, from a structural proteomics perspective." Curr Opin Chem Biol 8(1): 14-9.
- ^ Carrell, R. W. and D. A. Lomas (1997). "Conformational disease." Lancet 350(9071): 134-8.
External links
Categories:- Proteins
- Biochemistry
- Pharmacology
Wikimedia Foundation. 2010.